| |
In 1928, Menner established the genus Jaekelotodus for large sand tiger specimens from the Priabonian of Ukraine. Glikman (1964) erected the family Jaekelotodontidae for the teeth of Jaekelotodus (and in his opinion, for its ancestor Palaeohypotodus). Zhelezko & Kozlov (1999) included three species (with 12 named stages) to Jaekelotodus: J. londonensis ZHELEZKO, 1994, J. borystenicus Glikman 1964 and J. trigonalis (JAEKEL, 1895). Cappetta & Nolf (2005) included one additional species, J. robustus (LERICHE, 1921). Cappetta (1987) and Cappetta & Nolf (2005) rejected Glikman's creation of this new family. After examining numerous Jaekelotodus teeth, particularly from Kazakhstan, they concluded there was no particular characteristic(s) of the dental morphology or the overall dentition to distinguish them from other odontaspids; they included Jaekelotodus in Odontaspididae. Despite their assertions, Cappetta & Nolf provided little or no evidence for rejecting Glikman's family for these teeth. Lacking substantiated arguments, there appears no basis for rejecting Jaekelotodontidae as the family for these teeth. The Jaekelotodus evolution was interrupted due to climatic and other drastic global changes at the end of the Eocene (Zhelezko & Kozlov, 1999:.231-232).
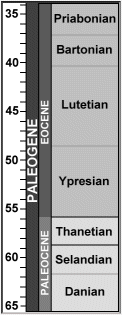
Two species have been reported in European faunas. Jaekelotodus robustus is found in the Thanetian of Belgium and France (Smith et al, 1999), more regularly in the Lower and Middle Eocene of Belgium (Leriche, 1905: 120) and in the Lower Eocene (London Clay) of Sheppey, UK (Casier, 1966).
Jaekelotodus trigonalis is rarely found in the Lutetian of Belgium (Leriche, 1905: 215) and France (Cappetta, 1987); it is relatively common in the French Bartonian (Dutheil, 1991) but never reaches the size of the Kazakh specimens. J. trigonalis is to the contrary, very common in the British Bartonian of Lee-on-the-Solent (Kemp, 1982)."
This species is also found in the former USSR (Jaekel 1895 & Glikman 1964) and has been reported from the Chesapeake Bay, USA (Ward & Wiest, 1990). In Western Europe, J. robustus teeth are reported from Thanetian to middle Eocene sediments, while J. trigonalis only appears in the middle Eocene. In the much diversified Ypresian fauna of Egem, only robustus occurs, being the least common 'sand tiger' at that location. In fact J. robustus is never abundant in the Western European fauna, but is much better represented in the Belgian Tertiary than trigonalis.
Characteristics of this genus' teeth include:
stoutly built teeth with massive roots, a strong lingual protuberance, clear nutrient groove and flat labial side
rather short, but broad (sometimes triangular) & completely smooth crown, often decreasing in width near the apex, a flat labial cusp face, medio-basally depressed, and a non-sigmoidal profile
one or two pairs of sharp and rather high cusplets.
sometimes lateral teeth can have small serrations where the cutting edge of the crown starts.
Jaekelotodus robustus (LERICHE, 1921)
This species has been described in detail by Leriche 1905 as Odontaspis crassidens AGASSIZ 1843 [In Agassiz 1843 pg. 292 as Lamna crassidens]. So first being considered as an Odontaspis, it was later attributed to the genus Hypotodus by Nolf (1986, 1988). Because of its dental morphology, this species can't be ascribed to the genus Odontaspis. Anterior Odontaspis teeth are slender, while robustus teeth are large and massive: in lateral teeth, the differences are even more apparent. The massive appearance of these teeth and the reduced size of the cusplets are enough to differentiate the species robustus from those of the genus Odontaspis. For similar reasons, they were attributed to the genus Jaekelotodus by Cappetta & Nolf (2005).
In general, the characteristics of isolated robustus teeth are similar to, but less pronounced than those of trigonalis. Anterior teeth possess a large, elongated crown (up to 36mm) with a smooth enameloid surface. The labial face overhangs the root and there is a medio-basal depression (less strong than seen in trigonalis). There is a complete cutting edge and a single pair of cusplets which are sharp and high. The root is massive and robust with a thick protuberance (more so in lower anteriors), a large nutrient groove and foramen. The root lobes are long, acutely splayed in the A1 position and becoming more obtuse distally.
Lateral teeth are also smooth. Upper laterals have a wide crown which is an elongated triangle in shape. The teeth are distally inclined, sometimes very strongly (more so than in trigonalis). Lower lateral crowns are more slender than their upper counterparts, being straight or only slightly inclined.
In lateral files, the distally inclined crown flattens labio-lingually. The cusplets become broader, diverging and are often double. The roots remain large with a distinct protuberance and nutrient groove; however, it is more apico-basally compressed than the anterior files and the well pronounced lobes are obtusely splayed.
Cappetta & Nolf (2005: p.247) write that in some cases, serrations may appear on the lower cutting edges of the crown. This couldn't be confirmed with our material. Their fifth tooth (plate 5, fig. 5 on p.264 ) should in the senior author's opinion be attributed to trigonalis rather than robustus.
Jaekelotodus trigonalis (JAEKEL, 1895)
The Jaekelotodus trigonalis dentition is mostly known from large (up to 40mm, more commonly 2.5 - 3 cm) isolated teeth and vertebra; however a partially articulated specimen (atypical and from a very old individual, figure 1), suggests a heavy-duty cutting dentition lacking the lower grasping teeth typical of odontaspids1. No articulated dentitions or associated tooth-sets have been published, and previous authors such as Cappetta (1987), Zhelezko & Kozlov (1999) and Cappetta & Nolf (2005)) have not tendered an opinion on a tooth formula. The accompanying figure (1) shows three files of lower anteriors plus seven lower lateral files. With a lack of detailed reporting on the dentition, it can only be said there are two or more upper anteriors (likely 3), three lower anteriors and seven or more files of lateral teeth in each quadrate. Cappetta & Nolf (2005) reported parasymphyseals not illustrated. Kemp (1994) figured an upper intermediate and lower parasymphyseals.
Anterior teeth are characterized by their straight and broad crowns, not very tall when compared with their massive and robust roots.
Upper anterior teeth are large, but less massive than the lowers. Upper anteriors have flatter cusps with more elongated crowns (sometimes triangular), while lowers have slightly lower, wider and thicker crowns with a stronger lingual protuberance.
The labial face has a strong medio-basal depression (V-shaped) and clearly overhangs the root. The enameloid is completely smooth. The cutting edge is complete, reaching the base of the cusp. There are two pairs of cusplets, a high and sharp principal pair and a tiny secondary. As opposed to the laterals, anterior teeth rarely possess serrations on the basal portion of the cutting-edge prior to the cusplets.
Our experience suggests that when viewed in profile, the first two significant lower anteriors may be lingually curved and the third, relatively straight. Only two upper anteriors have been referred to or illustrated in publications, leaving undetermined the upper anterior file count. It can be said that the profile of the mesial-most upper anterior is lingually curved, and the distal-most, erect.
The root lobes are long, acutely splayed in the A1 position and becoming more obtuse distally. They have a clear protuberance with a distinct nutrient groove and foramen.
Lateral teeth have smaller (slightly lower and narrower) crowns, getting broader at its base (triangular). Upper laterals are distally inclined, while lowers are straight or only slightly inclined. The labial crown face of lateral teeth overhangs the root, but with a much less marked medio-basal depression. The enameloid is completely smooth. The cutting edge is complete, often serrated basally and terminates in multiple cusplets (usually two). The mesial-most cusplet is the largest, broadly triangular and usually medially directed (towards the cusp). On occasion, both cusplets are merged forming one large, triangular cusplet. The lateral-most cusplet(s) are reduced and diminish in size laterally. Viewed in profile, the first few lateral files tend to be erect, becoming lingually curved in more distal file positions. The roots are still large with a distinct protuberance and nutrient groove; however it is more apico-basally compressed than the anterior files and the lobes obtusely splayed.
J. trigonalis can be distinguished from J. robustus by having anterior teeth with two pairs of cusplets (rather than one) and lateral teeth with higher and sharper cusplets. The nutrient groove is less marked than with J. robustus, never possess basal serrations and its anteriors have a slightly larger crown. Nolf (1988) included as differences between these species; robustus has a slightly convex labial crown face (profile view) and has a less angular labial crown base.
Jaekelotodus trigonalis is one of the scarcest species in the middle Eocene of Belgium. Only a very limited number of useful teeth have been found, making it difficult to draw firm conclusions. Differences occur when comparing these specimens with trigonalis from Kazakhstan. This can probably be explained by their different age (middle vs. upper Eocene) and / or different geological region. Our Belgian specimens are much smaller and the lowers are less robust (narrower crowns).
Remarks
This page includes what the authors consider possibly contradictory information. In looking at the articulated lower anteriors of J. trigonalis, there appears to be no stunted A1 (parasymphyseal sensu Cappetta & Nolf); in robustus, this tooth position is present. If viewed as a 'parasymphyseal' this could hardly be deemed as significant, however, if it is viewed as a stunted A1, then it would be. We follow Glikman, considering Jaekelotodus to be sufficiently unique to warrant its own family. With less conviction, we are following Cappetta and Nolf by including robustus in this genus.
Footnotes
| 1. |
The Hubbell specimen is unusually large and possibly deformed. Although it is useful for determining the number of file positions, it can be very misleading on typical crown design. |
Selected References
Agassiz, L. J. R. 1833-1844. Recherches sur les poissons fossiles. Text (5 vols; I., xlix+188 pp., II xii+310+366 pp., III viii+390 pp., IV xvi+296 pp., V xii+122+160 pp.) and Atlas (5 vols; I 10 pl., II., 149 pl., III 83 pl., IV, 61 pl., V, 91 pl.). Neuchâtel.
Cappetta, H. 1987. Chondrichthyes II. Mesozoic and Cenozoic Elasmobranchii. In: Handbook of Paleoichthyologie, vol. 3b, Gustav Fischer Verleg, Stuttgart, 193 pp.
Cappetta, H & Nolf, D, 2005. Revision de quelques Odontaspidae (Neoselachii: Lamniformes) du Paleocene et de l'Eocene du Bassin de la mer du Nord Bulletin de l'institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre 75: 237-266.
Casier, E. 1966. Faune ichthyologique du London Clay. Brit. Mus. Nat. Hist. ed., London, 496 pp.
Dutheil, D. B. (1991). A checklist of Neoselachii (Pisces, Chondrichthyes) from the Palaeogene of the Paris Basin, France. Tertiary Research 13(1): 27-36.
Glikman, L., 1964. Akuly paleogena i ikh stratigraficheskoe znachenie Akademii Nauk Soyuza Sovetskikh Sotsialisticheskikh Respublik, 1-228 Moscow.
Jaekel, O., 1895. Unter-Tertiäre Selachier aus Südrussland. Mémoires du Comité Géologique de Saint-Pétersbourg, 9: 19-35.
Kemp, D., 1982. Fossil sharks, rays and chimaeroids of the English Tertiary period, Gosport Museum.
Kemp, D., Kemp, L. and Ward, D., 1990. An Illustrated Guide to the British Middle Eocene Vertebrates. Ward, London, 59 pp.
Leriche, M., 1905. Les poissons tertiaires de la Belgique. II. Les poissons éocènes. Mém. Mus. Roy. Hist. Natur. Belg., 11(3): 49-228.
Leriche, M., 1921. Sur les restes de poissons remaniés dans le Néogène de la Belgique. Leur signification au point de vue de l'histoire géologique de la Belgique pendant le tertiaire supérieur. Bulletin de la Société belge de Géologie, 30: 115-120.
Menner, V. V. 1928. The Palaeogene sharks of Mangyschlak, Emba and from the east of Oural. Bulletin de la Société des Naturalistes de Moscou Section, Géologique vol. 6(3-4): 291-338. Moscow In Russian.
Smith, R., Smith, T. & Steurbaut, E. 1999. Les élasmobranches de la transition Paléocène-Eocène de Dormaal (Belgique) : implications biostratigraphiques et paléobiogéographiques. Bull. Soc. Geol. France, t.170 (3): 327-334.
Ward, D. J. and Wiest, R.L., 1990. A checklist of Paleocene and Eocene sharks and rays (Chondrichthyes) from the Pamunkey Group, Maryland and Virginia, USA. Tertiary Res., 12(2): 81-88.
Zhelezko, V.I., 1994. Sharks of family Jaekelotodontidae of European and middle Asian paleobiogeographic provinces. Bulletin Moskovian Society Naturalists, 69 (6): 47-62.
Zhelezko , V. I. & Kozlov, V. A. 1999. Elasmobranchii and Palaeogene biostratigraphy of Transurals and Central Asia. Materials on stratigraphy and Palaeontology of the Urals Vol. 3. Russian Academy of Sciences Urals Branch Uralian Regional Interdepartment Stratigraphical Comissian, Ekkaterinburg. 324 pp, 61 pls.
|