Isurus escheri (AGASSIZ, 1844)Lutz Andres ©2002 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
| Fig. 1: Upper A2 — Comparison Hastalis & Xiphodon: Delden Mbr, uppermost Miocene to lower Pliocene Langenboom/community of Mill / Holland, Escheri: 'postmiocene basisgrind/-laag', Lillo Frm., Doel near Antwerp / Belgium |
Temporal and geographical distribution
Isurus escheri appears in the middle to upper part of the Twistringer Formation of Germany (upper Middle Miocene) and according to van den Bosch et al. (1975) in the eastern Netherlands, "... in the Eibergen [lower - middle Late Miocene] for the first time." Any statements about the first appearance in Belgium are difficult to verify, because you have e.g. in Kallo, near Antwerp, the whole Miocene reworked as a 10 cm (~ 4 inch) thin layer -- the "postmiocene basisgrind/-laag" (post-Miocene basal conglomerate), between the Oligocene clay and Pliocene deposits. This species is present in that condensed Miocene layer, but there are both 'early' and 'typical' ones together (see Figs. 3 & 4) , as opposed to the Eibergen Member's fauna. "The teeth of Isurus escheri [in Eibergen Mbr] are not yet as 'typical' as they are in the [younger] Zenderen Mbr, the crenulations of the crown's cutting-edge are usually still rather indistinct. The impression exists that here, this species is still in an earlier evolutionary stage." (van den Bosch et al., 1975).
The first and last occurrence of Escheri in Denmark, France and Switzerland is currently unknown to the author, and deeper research into the literature and foreign collections is required. During the Late Miocene, I. escheri generally becomes a common species (van den Bosch et al., 1975) and the teeth represent a very typical form of this species (van den Bosch, 1969). The relationship of Hastalis and Escheri in the Eibergen Member (lower - middle Late Miocene) is 10:1 (van den Bosch, 1978 & 1980), and personal observations have shown that relative to Hastalis, the frequency of I. escheri in the Gramian (upper Late Miocene) is approximately 1:4, as observed while collecting at Kervenheim/Lower Rhine District/Nordrhein-Westfalen/Germany.
In Belgium, I. escheri becomes rare in lower Pliocene sediments (likewise Hastalis), and disappeared during the middle Pliocene (Sand of Oorderen). In contrast, the first appearance of Great White shark (Carcharodon carcharias) fossils are in the Delden Mbr (upper most Miocene - lower Pliocene, Netherlands), Sand of Kattendijk (lower Pliocene, Belgium) and Sand of Oorderen (middle - upper Middle Pliocene, Belgium).
 The accompanying Temporal Overview summarizes this transition geographically and stratigraphically over time.
The accompanying Temporal Overview summarizes this transition geographically and stratigraphically over time.
Odontological Characteristics
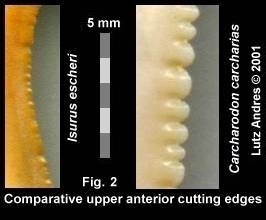 The cutting-edge of I. escheri teeth is crenulated, not weakly serrate. The strength of this crenulation, even on the largest specimens (e.g. 52 mm length), neither reach the development of a real serration, nor of the weak ones of some Pacific transitional teeth. There is a microstructural difference between a crenulated cutting-edge and one which is weakly serrate — comparable to the difference between emery paper and a saw. Similar to emery paper, the cutting-edge of Escheri teeth has more or less tiny grains which produce an irregulation. A serration however, has more or less clearly developed sharp points, the sum of which produce a saw-like cutting edge. In Escheri, the crenulated edge, as seen in Miocene specimens, appears to be a preliminary stage in the development of a fully serrate cutting-edge. However, until its disappearance from the fossil record some 10 million years later, there is no further changes to the design. In the Pacific 'transitionals' the development to a fully serrate cutting-edge proceeded, compared to Escheri, relatively fast.
The cutting-edge of I. escheri teeth is crenulated, not weakly serrate. The strength of this crenulation, even on the largest specimens (e.g. 52 mm length), neither reach the development of a real serration, nor of the weak ones of some Pacific transitional teeth. There is a microstructural difference between a crenulated cutting-edge and one which is weakly serrate — comparable to the difference between emery paper and a saw. Similar to emery paper, the cutting-edge of Escheri teeth has more or less tiny grains which produce an irregulation. A serration however, has more or less clearly developed sharp points, the sum of which produce a saw-like cutting edge. In Escheri, the crenulated edge, as seen in Miocene specimens, appears to be a preliminary stage in the development of a fully serrate cutting-edge. However, until its disappearance from the fossil record some 10 million years later, there is no further changes to the design. In the Pacific 'transitionals' the development to a fully serrate cutting-edge proceeded, compared to Escheri, relatively fast.
 |
| Fig. 3: Upper A2 — Early height=43, width=31, length=51 mm; 'postmiocene basisgrind/-laag', Lillo Frm., Kallo near Antwerp / Belgium |
The shape of the crown of upper anteriors is often much more slender and the cutting edge less rectilinear than those of Hastalis. The cutting edges don't extend in a straight line from the root to the tip; very often, they are much more irregular. However, the cutting edge of the 'early' ones is nearly identical to Hastalis suggesting a relationship between the two species. The root lobes of upper anteriors look more like those of the narrow-form Hastalis (= I. hastalis, more rounded) than those of the broad-form Hastalis (= I. xiphodon, more square).
 |
| Fig. 4: Upper A2 — Typical height=48, width=33, length=54 mm Delden Mbr., uppermost Miocene to lower Pliocene, Liessel / Holland. |
| Unlike the teeth of Hastalis, Xiphodon and the Great White, the upper third anterior (A3)2 of Escheri is not as triangular, being more hooked, like those of I. oxyrinchus (a.k.a. desori). |  |
| Fig. 5: Upper A3 height=28, width=22 mm; Uedem-layers, Upper Miocene/Gramium Kervenheim / Germany |
 |
| Fig. 6: Upper A3 variations ascribed to I. escheri A - Upper part of Langenfeldium (= lower Upper Miocene); Sittensen / G Möller collection B - Gramian (= upper Upper Miocene); Kervenheim / G; Andres collection C - Delden Mbr (upperst Miocene to lower Pliocene); Mill / NL; Morgenstern collection. (Click for enlargements of A and C.) |
| The lower anteriors generally have a much more I. oxyrinchus (a.k.a. desori)-look than a Hastalis-look. They are much more slender than those of Hastalis and/or Xiphodon, the crown sometimes shows a strong waist and the roots are very short and thick. This root gives these teeth a unique appearance that allows one to indentify them from a distance without examining the cutting edge. |  |
| Fig. 7: Lower A1 height=36, width=34 mm 'postmiocene basisgrind/-laag', Lillo Frm., Doel near Antwerp / Belgium |
 |
| Fig. 8: Lower A2 height=35 width=23 mm; 'postmiocene basisgrind/-laag', Lillo Frm.; Doel near Antwerp / Belgium |
Lateral teeth and those of juveniles/sub adults (and a few adults) often show distinct cusplets. One upper lateral tooth of Isurus escheri (30 mm length), which was found in the "Glimmerton" (mica clay) of the clay-pit "Sundern"/Twistringen near Bremen/ Niedersachsen/Germany, shows two to three cusplets on each side (see: http://home.t-online.de/home/rainer.meyer/twist.htm).
 |
| Fig. 9: Upper Lateral height=30, width=26, length=37 mm; Delden Mbr, uppermost Miocene to lower Pliocene Langenboom/community of Mill / Holland |
 |
| Fig. 10: Upper Lateral height=23, width=21, length=30 mm; Delden Mbr, uppermost Miocene to lower Pliocene Langenboom/community of Mill / Holland |
 |
| Fig. 11: Upper A1 - Juvenile height=27, width= 15, length=28 mm; Delden Mbr, uppermost Miocene to lower Pliocene Langenboom/community of Mill / Holland |
 |
| Fig. 12: Upper A2 - Juvenile height=25, width=16, length=28.5 mm; 'postmiocene basisgrind/-laag', Lillo Frm., Doel near Antwerp / Belgium |
The teeth of I. escheri never reach the size of those of Xiphodon. Based on published material and personal observations, Escheri uppers reach a length of 55 mm and lowers only 40 mm.
 |
| Fig. 13: Isurus escheri — Anterior files Upper A1-A3, L1 and Lower A1-A3 Not to scale and from different locations and horizons From the collections of Pepijn Morgenstern (uA3) & Lutz Andres |
In 1991, Hoepfner reported on the find of an Isurus escheri, including over 45 associated teeth and a few vertebrae, visible in the matrix. It was found in the clay layer ("Glimmerton") of the gravel-pit "Ohle"/Groß Pampau near Hamburg/Schleswig-Holstein/Germany by collectors in 1989. It is currently in the possession of the Museum of Natural History Luebeck and additional details remain unpublished.
Conclusions
 The temporal appearance in the Middle-Miocene and overall design of the 'early' Isurus escheri teeth allows one to presume that this species developed from Hastalis in the middle Middle Miocene. There is a noticeable onward evolution to the 'typical' Escheri design. Teeth of this species, the 'real' I. escheris, are found in Western Europe (Denmark, Netherlands, Belgium, France, Germany and Switzerland) only. Similar to its counterpart Isurus planus, which was obviously a Pacific species, I. escheri was an Eastern Atlantic species. This transition is summarized in the Escheri Evolution overview.
The temporal appearance in the Middle-Miocene and overall design of the 'early' Isurus escheri teeth allows one to presume that this species developed from Hastalis in the middle Middle Miocene. There is a noticeable onward evolution to the 'typical' Escheri design. Teeth of this species, the 'real' I. escheris, are found in Western Europe (Denmark, Netherlands, Belgium, France, Germany and Switzerland) only. Similar to its counterpart Isurus planus, which was obviously a Pacific species, I. escheri was an Eastern Atlantic species. This transition is summarized in the Escheri Evolution overview.
Why did it go extinct? It seems likely that the explanation is directly related to the evolution of the Great White shark. The Mega-toothed shark (Carcharocles megalodon), hunted large cetaceans (whales) and the next smaller ecological niche for a smaller predator, like the broad-form of Hastalis (= I. xiphodon), was to hunt small cetaceans (e.g. dolphins and porpoises) and pinnipeds (elephant seals and see lions). Flesh is easier to cut with serrate teeth and somewhere in the upper Miocene, e.g. in the Pisco Formation of Peru (de Muizon & de Vries, 1985) and/or in the Senhata Formation (Yabe, 2000), a few animals of a population of broad-toothed makos got weakly serrate teeth. They could feed more effectively and efficiently, therefore they could survive better and have more descendents. This characteristic was passed on to future descendants becoming ever more marked. The serrations become stronger over time and the result was the tooth-design represented in the extant Great White shark.3 The Great White shark developed in the Mio-Pliocene and spread out to be come a well suited circum-global predator. It prevailed over its own ancestor I. xiphodon and all other predators of nearly the same size and/or ecological niche (I. hastalis, I. planus and I. escheri). Presumably, the I. planus extinction occurred much earlier than that of I. escheri, because competition in the Pacific with the early Great White (Carcharodon sp.), was much earlier and more direct.
Therefore, I'm convinced that Isurus escheri is a side branch of the Isurus-lineage like the Great White, but dead ending — the Great White won the competition. Nature tested two prototypes of a broad-toothed mako and only one survived.
Acknowledgements
Special thanks to my good old friend Mike Möller (Kleve/Lower Rhine District / NRW / Germany), who was a source of inspiration and donated to me, more or less, teeth for my research. Further thanks to my other friends Renč Kindlimann (Aathal / Swiss) and Steven A. Alter (Florida / USA) for their helpful ideas, comments, etc. I must not forget Pepijn Morgenstern and Kris Blomme who opened their collections for this project, Dietmar Weber for his excellent artwork and Jim Bourdon for his assistance in editing this paper and preparing my images for presentation.
Review Comments
Both Mikael Siverson and David Ward reviewed the content (data not interpretative) of this page. They made several comments most of which have been incorporated within the text. The exceptions being Lutz's usage of the names xiphodon and desori (in accompanying Escheri Evolution chart) each of which they deemed to be nomen dubium. As noted in the introduction, the editor does not feel the abandonment of xiphodon as been sufficiently well argued and has not asked Lutz to make these changes. Although Mio-Pliocene desori teeth are now largely referred to as oxyrinchus, the true nature of Oligocene mako-like teeth is less clear, or at least poorly argued in print. The name desori was retained until better information is available.
Footnotes
| 1. |  The author asked Dietmar Weber to illustrate Isurus hastalis/ xiphodon. The reconstruction was based on facts like the tooth shape and size, probable diet/ecological niche, the guessed relationship to Isurus oxyrinchus (a.k.a. desori) & the Great White shark, and on the opinion and imagination of the author.
The author asked Dietmar Weber to illustrate Isurus hastalis/ xiphodon. The reconstruction was based on facts like the tooth shape and size, probable diet/ecological niche, the guessed relationship to Isurus oxyrinchus (a.k.a. desori) & the Great White shark, and on the opinion and imagination of the author. |
| 2. | By the way — there is an easy method to identify and orient the upper third anterior (in all Lamnid species) by looking at the shape of the root. Viewed basally, the tip of the distal root lobe is thinner and curved up, which causes a dent or depression on the lingual face of the root. In contrast, the mesial one is thicker and curved down or remains horizontal at the end (see: illustrations). This likely results from the distal lobe being positioned on the distal rise of the anterior hollow. |
| 3. | Stewart and Raschke (1999) examined corresponding teeth from 19 sites in the Capistrano Formation (Mio-Pliocene) of Orange County/California/USA and got a highly significant correlation (r = 0.925 — mathematical nearly 100%) between serration size and stratigraphy. They observed a measurable development of serration in the "Isurus hastalis - Carcharodon carcharias lineage" over time. |
References
| van den BOSCH, M. | |
| 1969. - Het Mioceen van Delden III. De Selachierfauna uit de miocene Afzettingen in het Twente-kanaal bij Delden. Publ. Natuurh. Genootsch. Limburg, 19, 1-2: 25-36, 70 figs. | |
| 1978. - On shark teeth and scales from the Netherlands and the Biostratigraphy of the Tertiary of the eastern part of the country. Meded.Werkgr.Tert.Kwart.Geol., vol. 15, nr. 4, p. 129-136. | |
| 1980. - Elasmobranch associations in the Tertiary and Quaternary deposits of the Netherlands (Vertebrata, Pisces), 2. Paleogene of the eastern and northern part of The Netherland, Neogene in the eastern part of The Netherland. Meded.Werkgr.Tert.Kwart.Geol., vol. 17, nr. 2, p. 65-70. | |
| van den BOSCH, M., CADEE, M.C. & JANSSEN, A.W., 1975. Lithostratigraphical and biostratigraphical subdivision of Tertiary deposits (Oligocene - Pliocene) in the Winterswijk-Almelo region (eastern part of the Netherlands). Scripta Geol. 29, p. 56-167. | |
| COOREMAN, P., 1997. La Belgique Cenozoique - Notice mineralogique et paleontologique. ASN. | |
| de CEUSTER, J., 1976. Stratigrafische Interpretatie van jong-cenozoische Afzettingen bij Rumst (Belgie, Province Antwerpen) en Beschrijving van de in een post-mioceen Basisgrind aangetroffen Vissenfauna, II. Systematische Beschrijvingen en Conclusies. Meded.Werkgr.Tert.Kwart.Geol., vol. 13, nr. 4, p. 119-172. | |
| HOEDEMAKERS, K., 1992. Fossilien aus dem Neogen von Belgien. Fossilien. 6/92, p. 351-357. | |
| HOEPFNER, G., 1991. Aus der Urzeit aufgetaucht: Wale und Haie - Bericht ueber Funde Luebecker Geschiebesammler aus dem Miozaen in Groß Pampau 1989. Der Geschiebesammler, vol. 24, nr. 3-4, p. 107-114. | |
| von der HOCHT, F., 1988. Die Knorpelfischfauna der Walfundstelle von Kervenheim. Natur am Niederrhein, 3,1, p. 13-24. | |
| in't HOUT, W., 1985. Haaietanden - en andere Tertiaire visresten uit Kallo (Belgie). gea, vol. 18, nr. 4, p. 125-160. | |
| in't HOUT, W. & OTTEMA, J., 1987. Fossilien uit het 4'e Havendok by Kallo, Belgie. Geologisch Museum Universiteit van Amsterdam. | |
| JANSSEN, A.W., van den BOSCH, M. & von der HOCHT, F., 1988. Age determination of a Tertiary deposit at Kervenheim in the Lower Rhine District, in connection with the find of a whale skeleton (F.R.G., Nordrhein-Westfalen). Rapport 124, Rijkmuseum van Geologie en Mineralogie, Leiden. [+ German translation] | |
| KLOSTERMAN, J., 1987. Ein fossiler Wal aus dem Tertiaer von Kervenheim. Natur am Niederrhein, 2,2, p. 43-54. | |
| KRUCKOW, T. | |
| 1960. - Obermiozaene Selachier von Hamburg-Langenfelde (Langenfelder Stufe). Verh.Ver.naturw.Heimatforsch.Hamburg, Bd. 34, p.55-61. | |
| 1965. - Die Elasmobranchier des tertiaeren Nordseebeckens im nordwestdeutschen Bereich. Senck.leth., nr. 46a, p.215-256. | |
| LINEAU, H.-W. | |
| 1987. - Haie und Rochen aus dem Sylter Ober-Miozaen. Fossilien von Sylt II, p. 19-75. | |
| 1992. - Fundumstaende von Chondrichthyer-Resten in miozaenen Glimmertonen des nordwestdeutschen Tertiaer-Beckens. Kaupia - Darmstaedter Beitraege zur Naturgeschichte, nr. 1, p. 71-87. | |
| MOTHS, H., 1998. Die Hai- und Rochenfauna aus dem Miozaen (Langenfeldium) von Groß Pampau. Der Geschiebesammler, vol. 31, nr. 2, p. 51-113. | |
| de MUIZON, C. & de VRIES, T.J., 1985. Geology and paleontology of late Cenozoic marine deposits in the Sacaco area (Peru). Geologische Rundschau, 74/3, p. 547-563. | |
| NOLF, D., 1986. Haaie- en Roggetanden uit het Tertiair van Belgie. Koninklijk Belgisch Instituut voor Natuurwetenschappen, Brussel. | |
| PURDY, R., SCHNEIDER, V., APPELGATE, S., MCLELLAN, J., MEYER, R. & SLAUGHTER, R., 2001. The Neogene Sharks, Rays, and Bony Fishes from Lee Creek Mine, Aurora, North Carolina. In: Geology and Paleontology of the Lee Creek Mine, North Carolina, III. C. E. Ray & D. J. Bohaska eds. Smithsonian Contributions to Paleobiology, No 90. Smithsonian Institution Press, Washington D.C. pp. 71-202. | |
| SIVERSON, M., 1999. A new large lamniform shark from the uppermost Gearle Siltstone (Cenomanian, Late Cretaceous) of Western Australia. Transactions of the Royal Society of Edinburgh: Earth Sciences, Vol 90, pp 49-66. | |
| STEWART, J.D. & RASCHKE, R., 1999. Correlation of stratigraphic position with Isurus - Carcharodon tooth serration size in the Capistrano Formation , and its implications for the ancestry of Carcharodon carcharias. Journal of Vertebrate Paleontology, 19(3), 78A. | |
| VERSCHUEREN, S., 1999. Fossiele Haaien, Roggen en Draakvissen van het Strand bij Cadzand en Nieuwvliet-Bad. In: Strandfossielen - van Cadzand en Nieuwvliet-Bad [Netherland]. geode, Nederlandse Geologische Vereniging Afdeling Amsterdam. | |
| YABE, H., 2000. Teeth of an extinct great white shark, Carcharodon sp., from the Neogene Senhata Formation, Miura Group, Chiba Prefecture, Japan. Tertiary Research, 201-4), p. 95-105. | |