Plinthicus cf stenodon COPE 1869
Like most Lee Creek visitors, I started as a surface collector -- walking the mine and picking up
what I "saw." Looking down at the tailings, Plinthicus teeth usually appeared as broken myliobatoid
teeth -- this proved to be an excellent example of "you find what you're looking for." When I first
saw an illustration in Cappetta (1987), I knew I'd been passing over many of these teeth; but looking
through my Lee Creek material, I managed to find a few examples. In subsequent visits to the mine,
I found and retained numerous examples of them.
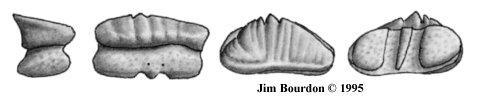
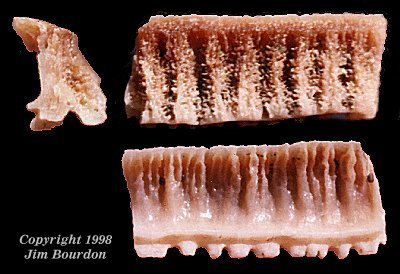
U-Type Teeth. The species appears to have had a rhinopterid-type dentition
with multiple, weakly-interlocking columns of teeth. Most striking in these teeth is the 45-degree
angle of recline seen in lateral profile. In occlusal view (figure 38, second from right), the hexagonal
myliobatoid shape can be seen. However, these teeth are labio-lingually compressed and very
high (relative to the other dimensions).
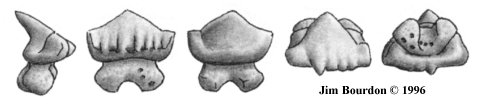
The most common are those illustrated in figure 5. Termed Type "U2", these are the laterally
shortened variation with about eight (range 7 to 9) root loblets. Next most common are the more
elongated teeth ("U3") with 12 (range 11 to 14) loblets. The propensity for breakage is unknown,
but the elongated teeth ("U4") with about 20 loblets appear less common. The specimen
illustrated in figure 6 is the only complete elongated specimen I've collected so far.
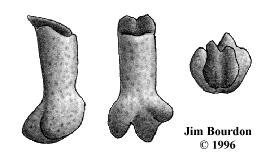
A mesio-distal slope of the occlusal face is pronounced only in some U2-type teeth, suggesting
a more lateral position. The only example of a true lateral tooth was provided by Becky Hyne
and is shown in figure 7.
 |
|
Fig 5. Type "U2" 96-JAB-TGA-174-B, width = 9.3 mm |
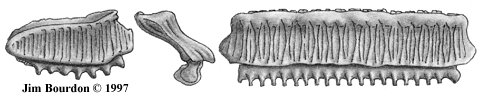 |
Plinthicus stenodon
Fig 7. (left) Type "U6" 9X-BH-TGA-61, width = 12.0 mm
Fig 6. (right) Type "U4" 96-JAB-TGA-174-E, width = 20.0 mm |