 |
|
|
|
| PUNGO RIVER FM. (Miocene) The warm waters of the Miocene were home to sharks and rays that are no longer reported from the today's North Carolina waters; many of the smaller gray sharks are now found along the Indian and western Pacific Oceans. Reported and unreported species include (updated species pages are notated by a linked image): |
 |
C. brachyurus (GÜNTHER, 1870) -- Copper shark
Abundant in Pungo River
Confidence: Very high | Upper reconstruction conforms with Recent tooth-set
Extant: Circumglobal, subtropical
Purdy et al (2001): Pungo units 1-5. |
 |
C. falciformis (BIBRON, 1839) -- Silky shark
Common in Pungo River
Confidence: Very high | Upper reconstruction conforms with Recent tooth-set
Extant: Circumglobal, temperate / subtropical
Purdy et al (2001): Pungo units 2-4. |
 |
C. cf isodon (VALENCIENNES, 1839) -- Finetooth shark
Scarce in Lower Pungo River tailings
Confidence: high | upper reconstruction conforms with Recent tooth-set. lowers cloaked by C. macloti.
Extant: Gulf of Mexico
Purdy et al (2001) do not include this tooth-design. |
| |
C. leucas (VALENCIENNES, 1839) -- Bull shark
Uncommon in the Yorktown, (smaller & less common in the PR).
Confidence: medium-high | Awaits tooth-set reconstruction
Extant: Circumglobal, subtropical
Purdy et al (2001) rare in PR units 1-5, common in YT units 1-3. |
|
C. cf limbatus (VALENCIENNES, 1839) -- Blacktip shark.
Uncommon Pungo River tailings
Confidence: low-medium | based on isolated tooth-positions
Extant: Circumglobal, subtropical
Purdy et al (2001) do not include this tooth-design. |
 |
C. macloti (MÜLLER & HENLE, 1839) -- Hardnose shark.
Abundant in Pungo River sands
Confidence: Very high | Upper reconstruction conforms with Recent tooth-set, lower teeth well represented
Extant: Indian Ocean and western Pacific
Purdy et al (2001): PR Units 1-5 & YT (unit 1, ?redeposit) |
 |
C. perezii (POEY, 1876) -- Caribbean Reef Shark
Abundant in Pungo River, common in Yorktown
Confidence: Very high | Upper reconstruction conforms with Recent tooth-set, lower teeth represented
Extant: Western Atlantic, subtropical
Purdy et al (2001): Pungo River units 3-5 and Yorktown Unit 1 |
 |
Carcharhinus plumbeus (NARDO, 1827) -- Sandbar shark
Uncommon in Pungo River, common in Yorktown
Confidence: High | Upper reconstruction conforms with Recent tooth-set
Extant: Scattered circumglobal, subtropical
Purdy et al (2001): Pungo River units 4,5); Yorktown unit 1 |
| Additional tooth-designs that could not be attributed to the above taxa are present in Pungo River tailings (a project for the future). |
| YORKTOWN FM. (Pliocene) The cooler waters of the Pliocene were home to sharks and rays that currently inhabit today's North Carolina waters; |
|
C. cf galapagensis
(SNODGRASS & HELLER, 1905) Galapagos shark
Relatively rare in the Yorktown and possibly from the Pungo River.
Confidence: Low-Med | Represented by a few isolated teeth and based on tooth-design only
Extant: Circumglobal, Tropical-subtropical
Purdy et al (2001) do not include this species in the fauna. |
| |
C. leucas (VALENCIENNES, 1839) -- Bull shark
Uncommon in the Yorktown, (smaller & less common in the PR).
Confidence: medium-high | Awaits tooth-set reconstruction
Extant: Circumglobal, subtropical
Purdy et al (2001) rare in PR units 1-5, common in YT units 1-3. |
| |
C. cf longimanus (POEY, 1861) -- Oceanic Whitetip shark
Unommon in the Yorktown.
Confidence: low-med | Awaits verification
Extant: Circumglobal, temperate-subtropical
Purdy et al (2001) assigned this design to C. leucas. |
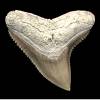 |
C. obscurus (LeSUEUR, 1818) -- Dusky shark
Most abundant gray shark in the Yorktown.
Confidence: high | Upper reconstruction conforms with Recent tooth-set, lower teeth represented.
Extant: Circumglobal, temperate-subtropical
Purdy et al (2001) reported from Yorktown units 1-3. |
|
|
C. perezii (POEY, 1876) -- Caribbean Reef Shark
Abundant in Pungo River, common in Yorktown
Confidence: Very high | Upper reconstruction conforms with Recent tooth-set, lower teeth represented
Extant: Western Atlantic, subtropical
Purdy et al (2001): Pungo River units 3-5 and Yorktown Unit 1 |
|
|
Carcharhinus plumbeus (NARDO, 1827) -- Sandbar shark
Uncommon in Pungo River, common in Yorktown
Confidence: High | Upper reconstruction conforms with Recent tooth-set
Extant: Scattered circumglobal, subtropical
Purdy et al (2001): Pungo River units 4,5); Yorktown unit 1 |
| Additional tooth-designs that could not be attributed to the above taxa are present in Yorktown tailings. |