Cappetta (1987) includes two extinct species, G. delfortriei DAIMERIES 1889 from the Miocene of Europe and Africa and G. malembeense DARTEVELLE & CASIER 1943 from Miocene of West Africa.
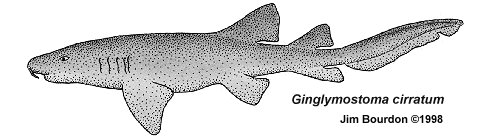
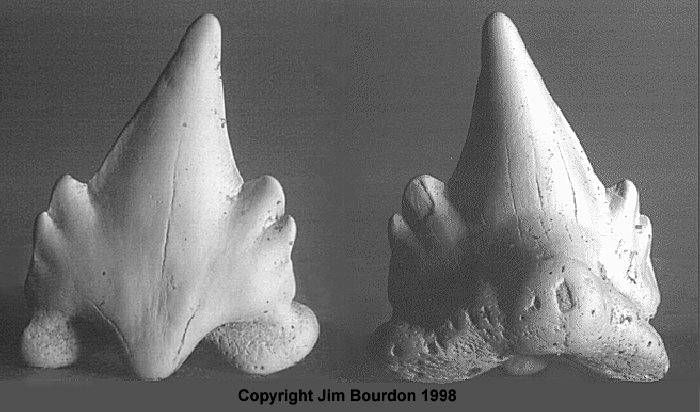
Ginglymostoma sp. Tailings from the mine in Aurora rarely yield a Nurse shark teeth. Those specimens seen by the author have all been from DPW material, thought to be of Pungo River Fm (Lower Miocene) origin. Most of these teeth have root damage, which would question the validity of any identification. However, the accompanying illustration is clearly a nurse shark. The author is of the opinion that these teeth likely represent the extant species G. cirratum
Selected References
Cappetta, H., 1987. Chondrichthyes II. Mesozoic and Cenozoic Elasmobranchii. In: Handbook of Paleoichthyologie, vol. 3b, Gustav Fischer Verleg, Stuttgart, 193 pp.
Cappetta, H. & Case, G., 1975. Contribution à l'étude des sélaciens du groupe Monmouth (Campanien - Maestrichtian) du New Jersey. Palaeontographica Abteilung A, 151:1-46.
Case, G., 1994. Fossil Fish Remains fron the Late Paleocene Tuscahoma and Early Eocene Bashi Formations of Meridian, Lauderdale County, Mississippi. Palaeontographica Abt. A, 230, pp 97-138.
Case, G. R., and H. Cappetta. 1997. A new selachian fauna from the late Maastrichtian of Texas. Muünchener Geowissenschaften Abhandungen 34:131-189.
Compagno, L.J., 1999. Checklist of living elasmobranchs. In Sharks, skates, and Rays - The Biology of Elasmobranch Fishes. Hamlett, W. C. ed. John Hopkins University Press, Baltimore MD, USA. pp 471-498.
Hamm, S.A. and D.J. Cicimurri, 2011. Early Coniacian (Late Cretaceous) selachian fauna from the basal Atco Formation, Lower Austin Group, north central Texas; Paludicola [Rochester Institute of Vertebrate Paleontology] 8(3):107-127.
Hartstein, E., Decina, L. and Keil, R., 1999. A Late Cretaceous (Severn Formation) Vertebrate Assemblage from Bowie, Maryland. The Mosasaur, 6:17-23.
Kent, B., 1994. Fossil Sharks of the Chesapeake Region. Egan Rees & Boyer, Maryland. 146 pp
Kent, B. 1999. Sharks from the Fisher/Sullivan Site. In: Weems, R. & Grimsley, G., Early Eocene Vertebrates and Plants from the Fisher/Sullivan Site (Nanjemoy Formation) Stafford County, Virginia. Virginia Division of Mineral Resources, Pub 152: 11-37.
Leriche, M., 1942. Contribution à l'étude des faunes ichthyologiques marines des terrains Tertiaires de la Plaine Côtière Atlantique et du centre des Etats Unis. Mémoire de la Société Géologique de France, Paris, new series, 43:1-111.
Müller, A. 1999. Ichthyofaunen aus dem atlantischen Tertiär der USA. Leipziger Geowissenschafteb, Leipzig, 9/10: 1-360.
Noubhani, A and Cappetta, H., 1997. Les Orectolobiformes, Carcharhiniformes et Myliobatiformes des Bassins à phosphate du Moroc (Maastrichtien-Lutétien basal)., PalaeoIchthyologica 8, München. 327 pp
Schwimmer, D., 1986. Late Cretaceous fossils from the Blufftown Formation (Campanian) in western Georgia. The Mosasaur, III:109-119.
Ward, D. and Wiest, R., 1990. A checklist of Paleocene and Eocene sharks and rays (Chondrichthyes) from the Pamunkey Group, Maryland and Virginia, USA. Tertiary Res., 12(2) p 81-88.
Welton, B. and Farish, R., 1993. The Collector's Guide to Fossil Sharks and Rays from the Cretaceous of Texas. Before Time, Texas. 204 pp.

{kind=link}