| |
Editor's note:
If there are any teeth that Paleogene collectors tend to lump, it is those of the sand tiger-design. As a group (odontaspid, jaekelotodontid & mitsukurinid in this webpage), they can be readily differentiated from other families, but ascribing these teeth to a specific taxon is often problematic. This identification problem is not limited to the amateur; the professionals have been neither clear nor consistent when attempting to identify and classify these teeth. Cappetta (1987) attempted to bring a semblance of order to this problem as did Ward (1988) and Kent (1994) but inconsistencies continued. Cappetta & Nolf 2005 is the most recent attempt to bring clarity to portions of this group — in this case, Paleogene sand tigers of Western Europe.
As this website is focused on the Western Atlantic, the Cappetta & Nolf publication provided four major challenges.
An accurate translation into English,
A brief discussion of the validity of the presented arguments,
Verification that the names ascribed to the European taxa are the same as their North American counterparts, and
Providing some value to readers from other areas.
These requirements required a joint effort. Pieter De Schutter joined the team to provide translation skills and a European perspective to the taxa. Steve Cunningham is a highly respected researcher of North American sand tigers and particularly knowlegeable in Striatolamia teeth. Together we hope to provide sufficient insight as to whether or not hundreds of thousands of specimens should be re-labeled. This paper will also serve as the basis for identification changes within elasmo.com.
An Overview
Cappetta & Nolf 2005 (C&N) is an ambitious attempt to bring order to the sand tigers of Western Europe. It not only addresses regional authors but ventures into the sometimes perplexing world of the Russian authors. Taxa are synonymized, genera erected or resurrected, and species moved and/or renamed. The text provides details on tooth-design and is supplemented with illustrations (usually single perspective) of various tooth positions for eight of the taxa. An exhaustive list of described taxa is included.
Jaekelotodontidae GLIKMAN 1964
With limited familiarity of Jaekelotodus trigonalis teeth (not common in the North American fauna), two of the authors (JB & SC) were unable to express any strong 'hands-on' experience for the genus Jaekelotodus. Referring to Zhelezko & Kozlov (1999) which included numerous figures of the species Jaekelotodus trigonalis, a general 'feel' for the species could be established. Zhelezko & Kozlov lumped anterior teeth without designation as to uppers & lowers (or provided lateral profile illustrations), so it could only be hypothesized that at least five (upper & lower combined) file-positions were represented. Zhelezko & Kozlov did distinguish the laterals; the important feature being that the lower laterals had relatively low, broad, triangular cusps. This feature is neither present in Carcharias nor Odontaspis, so it is understandable that Glikman (1964) perceived a different family and C&N a different genus.
Neither of these authors provided file-design details nor a tooth-set, so their observations cannot be properly judged or critiqued. Elasmo.com will accept Jaekelotodus as a distinct genus, but finds no evidence (valid arguments) in C&N for rejecting Glikman's assignment of J. trigonalis to its own family. The primary basis for this argument appears to be that:
| |
Le genre Jaekelotodus possède en particulier des dents parasymphysaires, tout comme les Carcharias ou les Odontaspis, et aucune particularité dentaire ne permet de les séparer des Odontaspidae. Pour ces raisons, nous continuons à considérer la famille des Jaekelotodontidae comme synonymes des Odontaspididae (voir Cappetta, 1987: 86).
In particular (and not some) Jaekelotodus have parasymphyseal teeth, just like Carcharias or Odontaspis, and no dental characteristic makes it possible to distinguish them from Odontaspididae. For these reasons, we continue to consider the family of Jaekelotodontidae synonymous with Odontaspididae.) |
|
Cappetta & Nolf appear to have ignored the robust root, low & wide cusp, and occasionally serrate cutting edge in favor of a parasymphyseal they neither described/illustrated nor tendered evidence of. Such certainty for the presence of these stunted teeth would suggest an articulated tooth-set not referred to. It is not understood by this webpage's authors how species can be assigned or re-assigned between taxa without an understanding of the tooth-set design.
Jaekelotodus MENNER 1928
C&N broadly defines this taxa and goes on to specifically discuss two species. Arguments are provided by the authors to separate these teeth from those of Odontaspis and assign them to their own genus. They rejected Glikman's (1964) arguments that this genus should be ascribed to a new family (Jaekelotodontidae) and retained it within Odontaspidae. They discussed two species which can be differentiated based on cusplets (trigonalis' anteriors having two pairs and those of the laterals being higher) and the nutrient groove (which is stronger in robustus).
Jaekelotodus robustus (LERICHE, 1921). Cappetta & Nolf extensively discussed their choice of this specific name. They went on to describe and illustrate these teeth and refer the reader, for a detailed description, to Leriche 1905.
They noted that over the years, teeth of this design had been attributed to:
Leriche (1905, 06) - Odontaspis crassidens
Leriche (1921, 31, 46, 59, 66) - Odontaspis robusta
Nolf (1986, 88) - Hypotodus robustus
Kemp et al (1990) - Jaekelotodus trigonalis
Teeth of this design have been reported from the Eocene of the Western Atlantic as Carcharias robusta (Kent 1994). De Schutter notes that in Belgium, they followed Nolf (1988) and preferred Hypotodus robustus.
Jaekelotodus trigonalis (JAEKEL, 1895). C&N provides descriptive text and illustrations for this species. Oddly, the illustrated specimens are from Kazakhstan.
They note that this species has been included in various genera:
Jaekel (1895), Nolf (1986, 88) - Hypotodus trigonalis
Jaekel (1899) - Otodus trigonalis
Menner (1928), Cappetta (1987) and Kemp et al (1990) - Jaekelotodus trigonalis
Casier (1966) - Odontaspis trigonalis
Ward & Wiest (1990) and Kent (1994) have reported Jaekelotodus trigonalis
from the Eocene of the Western Atlantic.
Mitsukurinidae JORDAN 1898
Cappetta and Nolf did not directly address the goblin sharks. In Cappetta (1987), Striatolamia GLIKMAN 1964 was ascribed to Odontaspididae; only the extinct taxa Anomotodon ARAMBOURG 1952 and Scapanorhynchus WOODWARD 1889 were included in Mitsukurinidae. In this new paper, they claimed to follow Siverson (1992, possibly intending to refer to 1995) and include Striatolamia as a mitsukurinid. In the Paleogene sediments of the Chesapeake Bay region, this simple reassignment takes 60-70% of the sand tiger group teeth out of consideration. Fortunately, Cunningham (2000) provides an extensive study of Striatolamia, which can in most cases, help differentiate these teeth from other sand tigers. Apparently the authors overlooked the significance of these observations.
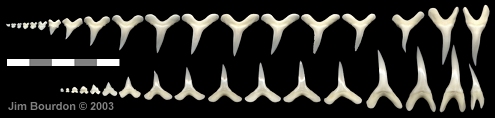 |
Fig. 1 - Mitsukurina owstoni adult dentition
extant taxon; Hubbell collection |
Despite the C&N assertions, this webpage's authors are not convinced of the assignment of these Striatolamia teeth to Mitsukurinidae. Based on these observations, elasmo.com will continue to include Striatolamia as an odontaspid.
Odontaspididae MÜLLER & HENLE 1839
In the wake of the Glikman Revolution, Cappetta (1987) included six Paleogene genera as odontaspids: Hypotodus JAEKEL 1895, Jaekelotodus MENNER 1928, Odontaspis AGASSIZ 1838, Palaeohypotodus Glikman 1964, Striatolamia Glikman 1964 and Synodontaspis WHITE 1931. As summarized in the preceding link, Compagno was successful in getting the ICZN to accept Carcharias RAFINESQUE, 1810 (in lieu of Synodontaspis). As mentioned earlier, Siverson (1995) moved Anomotodon (and by perceived association, Striatolamia) to the mitsukurinids, leaving five genera. C&N increases this collection of Western European1 genera to nine by including Glueckmanotodus ZHELEZKO & KOZLOV 1999 and erecting three new genera. Of these genera, those with well known teeth can be broadly placed into groups that correspond with the extant taxa Carcharias and Odontaspis.
Carcharias Affinity
The extant taxon, Carcharias taurus, has a regionalized, yet circumglobal distribution in warm temperate to tropical coastal waters. Its dentition can be summarized as having a tearing-design, a smooth lingual cusp face with:
3 upper anterior teeth
1-4 intermediate teeth
7 anterio-lateral teeth, and
6 or more reduced posterio-lateral teeth
a stunted A1 plus 3 significant lower anterior teeth (A2-A4)
6 anterio-lateral teeth, and
7 or more reduced posterio-lateral teeth.
Tooth-sets (genera) included in this group tend to follow the Carcharias dentition-design.
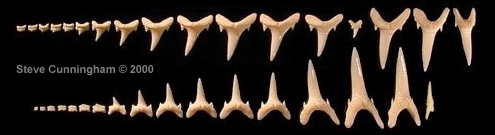 |
Fig. 2 - Carcharias taurus adult dentition
extant taxon; Cunningham collection |
Brachycarcharias nov. gen.
Cappetta & Nolf erected this genus for a sand tiger tooth-design long bounced between numerous genera and even families. They included two valid species within the scope of this paper, B. lerichei and B. atlasi.
Brachycarcharias lerichei (CASIER, 1946).
The authors provided a good description and illustrations of the adult teeth. Despite assigning these teeth to Odontaspidae they proposed no dental formula for the anterior teeth; they did note however, that the teeth are closest to the Carcharias tooth-design.
Teeth synonymized with or moved (all or in part) to B. lerichei include:
Woodward (1899), Leriche (1905, 06, 08, 22) - Lamna vincenti
Casier (1946) - Lamna lerichei
Arambourg (1952) - Odontaspis vincenti
Nolf (1970) - Odontaspis substriata mut. atlasi
Nolf (1972) - Odontaspis atlasi
Taverne & Nolf (1978) - Odontaspis atlasi
Nolf (1986, 88, 90) - Lamna lerichei
Cappetta (1987)2 - Lamna lerichei
Kent (1994)2 - Cretolamna lerichei
Baut & Genault (1995) - "Carcharias" vincenti
elasmo.com (1998)2 - Serratolamna lerichei
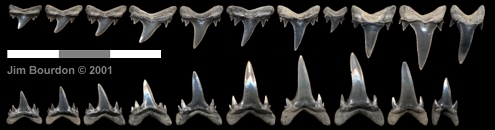 |
Fig. 3 - Brachycarcharias lerichei representative tooth-set
Juvenile or young adult; Nanjemoy Fm (Ypresian), Virginia; Bourdon collection |
In 2000-01, one webpage author (JB) employed the Cunningham (2000) horizontal dentition methodology to teeth of lerichei-design from the Ypresian of Virginia. It was concluded the overall design was odontaspid not lamnid (ref above representative tooth-set). The C&N conclusions appear valid to the authors of this review.
Carcharias RAFINESQUE 1810
Extinct sharks with a dentition and tooth-design of the extant taxon can be clearly traced back to the Oligocene (C. acutissima). In the appendix, C&N lists some 25 fossil species ascribed to Carcharias and challenges/reassigns most of them. None of the remaining C&N taxa have been reported from Europe. The authors argue that this taxon is represented in the Paleogene of Europe by Carcharias sp only and that other taxa (from that region) should be abandoned or ascribed to different genera.
Ward (1988) synonymized multiple sand tiger taxa into "Carcharias hopei". A major thrust of C&N is to demonstrate that this was an artificial genus composed of the teeth of other taxa. Unfortunately, the C&N arguments were largely directed towards naming conventions (rules, precedence, holotypes, neotypes, etc.) and not to the dentition's design.
Carcharias sp nov sp.
Cappetta & Nolf recognized a single Paleogene taxon (striated lingual face) which they viewed as having been overlooked by most previous authors. They described in text and provided illustrations for this new species which they deemed comparable to C. acutissima and therefore the oldest example of the extant C. taurus lineage.
They cited the few examples of this tooth-design previously reported.
Nolf (1986, 88) - Synodontaspis hopei
Kemp et al (1990) - Carcharias acutissima
The Belgian co-author for this webpage (De Schutter) notes that specimens of this Carcharias sp tooth-design have frequently been ascribed to teretidens and "hopei" in Europe. The similarity of these teeth to those of the Rupelian C. acutissima had been noted during the creation of the Belgiansharkteeth website and obviously to Kemp et al (1990). Combining this familiarity with the specimens and the details provided in C&N, this appears to be a valid taxon of the C. acutissima-taurus lineage.
Those unfamiliar with this fauna may feel poorly served by the cursory nature of the description and illustrations. Also perplexing is the lack of a detailed comparison with C. acutissima. They point out that it is similar but reject out of hand the Kemp et al determination.
Glueckmanotodus ZHELEZKO & KOZLOV 1999
Zhelezko & Kozlov erected this genus based on Thanetian specimens from Belgium and went on to describe two additional species (Selandian & Danian) from Central Asia. In the appendix, C&N draws no firm conclusions on these other teeth deeming them 'difficult to evaluate'.
Glueckmanotodus heinzelini (CASIER, 1967). C&N recognizes Casier's species and the subsequent assignment to Glueckmanotodus . The authors described these teeth and contrasted them with the Hypotodus verticalis tooth-design. Illustrations are provided to supplement these comparisons.
The authors noted that over the years, teeth of this design had been attributed to numerous genera and/or species:
Gurr (1962) - Odontaspis robusta and Lamna verticalis
Casier (1967) - Odontaspis heinzelini and O. hopei
Nolf 1986 - Hypotodus heinzelini
Ward (1988) - Carcharias hopei
Nolf 1988 - Hypotodus heinzelini
Baut & Genault (1995) - Jaekelotodus heinzelini, Palaeohypotodus rutoti and Carcharias hopei
At the time of this writing (Feb 2006), Glueckmanotodus had not been reported from the Western Atlantic; this may change based on the abandonment of Carcharias hopei (sensu Ward 1988). G. heinzelini is regularly found in the middle Thanetian of France (Moreau & Mathis 2000).
Hypotodus JAEKEL, 1895
Potentially the most controversial of the C&N conclusions is the rejection of Carcharias hopei as proposed by Ward (1988) (both genus and species) and reassigning many of these teeth (specifically the diagnostic teeth) to Hypotodus verticalis. Based on the appendix, C&N concludes that there is a single valid taxon; however, the authors suggest that further study may reveal that Arambourg's (1952) Odontaspis hopei subsp. atlantica may be referable to Hypotodus atlanticus (ARAMBOURG, 1952).
Hypotodus verticalis (AGASSIZ 1843)
The specific name to be employed for these teeth is more complex. C&N provides multiple reasons for rejecting C. hopei in favor of verticalis. Because Agassiz's (1844) illustrations of hopei teeth included more than one taxa (as C&N notes) one can only conclude, as Cappetta & Nolf did, that verticalis is a more appropriate name for the this species and its teeth.
Although incorporating a great deal of text directed to the matter of name (genus & species), C&N provides feeble detail on tooth and dentition-design. It only includes illustrations of a single anterior tooth-position and actually provides little or no evidence that these teeth should be ascribed to a genus other than Carcharias.
They noted that this species had been previously included in various genera:
Agassiz (1843) - ? Lamna hopei, L. verticalis
Winkler (1874b) - Otodus vincenti
Leriche (1908) - Lamna verticalis
Casier (1946) - Odontaspis hopei, O. verticalis
Nolf (1986, 88) - Syndontaspis hopei, Hypotodus verticalis
Cappetta (1987) - Syndontaspis hopei2, Hypotodus verticalis
Ward (1988), Kemp et al (1990) - Carcharias hopei
 |
Fig. 4 - Hypotodus verticalis artificial tooth-set
Juvenile or young adult; Nanjemoy Fm (Ypresian), Virginia; Bourdon collection
Reconstruction based on Cunningham (unpublished); asterisk: see footnote 3 |
Earlier work by Cunningham (unpublished, 2000) demonstrated that a significant upper A1 tooth-position is lacking in this taxon, clearly differentiating it from Carcharias. (The senior author has replicated the Cunningham observations for this webpage.) Despite the poor arguments and descriptions provided by Cappetta & Nolf, the assignment of these teeth to a different genus, in this case Hypotodus appears valid. To supplement their observations, we propose the above tooth-set.
Orpodon nov. gen.
Cappetta & Nolf erected a new genus for small sand tiger teeth from Western Europe which have over-sized (high, triangular and wide relative to the cusp) cusplets, smooth lingual crown face and occasionally folds on the lower labial face.
Orpodon heersensis (HERMAN, 1972). Cappetta & Nolf recognized a single species with this tooth-design, and found references to it under other names:
Winkler 1874a - Otodus parvus
Herman 1972 - Odontaspis heersensis
Herman 1977 - Jaekelotodus heersensis
This tooth-design has not been reported from the Western Atlantic.
Striatolamia GLIKMAN, 19642
As noted above, Striatolamia was not included as an odontaspid in C&N. Cunningham (2000) reconstructed a Carcharias-like tooth-set based on several hundred Striatolamia macrota teeth from the Nanjemoy Fm (Ypresian, Virginia). In the intervening years, increasing the sample to thousands of teeth only served to re-enforce these earlier conclusions (unpublished).
The senior webpage author had also collected this Potapaco Mbr, Nanjemoy Fm locale and attempted to repeat the Cunningham (2000) conclusions for this webpage. Using his 'horizontal dentition' methodology, the results were successfully duplicated. The similarity of the tooth-set design (lateral file-position counts and tooth-design) strongly supports an odontaspid dentition of the Carcharias-type.
 |
Fig. 5 - Striatolamia macrota artificial tooth-set
Juvenile or young adult; Nanjemoy Fm (Ypresian), Virginia; Bourdon collection
Reconstruction based on Cunningham (2000); asterisk: see footnote 3 |
At least two species have been reported from both sides of the Atlantic, Striatolamia macrota (AGASSIZ 1843) and S. striata (WINKLER, 1874a).
Sylvestrilamia nov. gen.
Cappetta & Nolf erected this genus for striated sand tiger teeth in which,
the anteriors have a straight cusp which lacks a strong sigmoidal profile.
The crown has a complete cutting edge & a pair of short pointed cusplets and the roots are thick & well splayed,
the lower laterals are upright and the uppers inclined, each with low rather broad cusplets.
They mentioned no other species associated with this genus, provided no details as to the tooth-set design, or made any suggestions as to its affinities (Carcharias vs Odontaspis). Lacking supporting illustrations and comparative details, the key diagnostic features for differentiating this genus from other odontaspids is unclear.
They noted that identification is difficult and largely based on the morphology of the cusplets.
Sylvestrilamia teretidens (WHITE, 1931). C&N provides a brief description and illustrations that include only a single anterior and upper tooth position. They referred the reader to White (1931) for additional details.
This tooth-design has been generally recognized over the years and attributed to teretidens:
White (1931), Casier (1946) - Otodus rutoti
Nolf (1986, 1988) - Synodontaspis teretidens
Cappetta (1987)2 - Synodontaspis teretidens
Kent (1994, 1999)2 - Carcharias teretidens
Teeth of this design have been reported from the Paleogene of the Western Atlantic (Kent 1994 & 1999).
Odontaspis Affinity
The extant taxon, Odontaspis ferox, has a spotty, yet circumglobal distribution in warm temperate to tropical coastal waters. Its dentition can be summarized as having a tearing-design, a generally smooth lingual cusp face with:
3 upper anterior teeth, A1 stunted
multiple intermediate teeth
20 or so upper lateral teeth with no clear break differentiating posteriors
a small A1 plus 3 significant lower anterior teeth (A2-A4)
15 or so lower laterals, with no clear break differentiating posteriors
The Odontaspis noronhai dentition is similar but the lateral teeth are broader and less numerous. In addition, a stunted upper A4 may be present, there are less or no intermediate teeth and the lower jaw may have multiple files of parasymphyseal teeth.
Tooth-sets (genera) included in this group tend to follow the Odontaspis dentition-design.
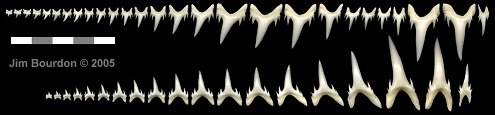 |
Fig. 6 - Odontaspis ferox adult dentition
extant taxon; Hubbell collection |
Odontaspis AGASSIZ 1838
Teeth very similar to those of the extant taxon, Odontaspis ferox RISSO 1810, are easily traceable to the Paleogene on both sides of the North Atlantic. The C&N appendix includes five species, three of which were questioned by the authors. Only the European species was included.
Odontaspis winkleri LERICHE, 1905.
C&N provides a short description of these teeth and no illustrations, referring the reader to Leriche (1905, 1906). Since first described by Leriche, succeeding (European) authors have been consistent in ascribing these teeth to O. winkleri.
Because teeth of the Odontaspis-design are common and readily recognized, there have been no (published) attempts to reconstruct this dentition; the unanimity of opinion may conceal the caveat of multiple species.
Palaeohypotodus GLIKMAN, 1964
Cappetta & Nolf have rejected or synonymized all Paleogene species attributed to this genus leaving only a single taxon - P. rutoti. they noted that these teeth can be distinguished from Odontaspis by the basio-labial crenulations and complete cutting edge.
Palaeohypotodus rutoti (WINKLER, 1874b). C&N provides little descriptive detail and no illustrations for this taxon, referring the reader to Leriche 1902, 1905, 1906 and 1951.
This tooth-design has been generally recognized over the years and attributed to P. rutoti, its generic assignment however has bounced around -- the authors note:
Winkler (1874b), Vincent (1876) - Otodus rutoti
Woodward (1891) - Odontaspis houzteaui
Leriche (1902, 1951), White (1931), Herman (1972) - Odontaspis rutoti
Glikman (1964), Cappetta (1987), Nolf (1988) - Palaeohypotodus rutoti
Palaeohypotodus rutoti has been reported from the Palaeocene of the Western Atlantic (Ward & Wiest 1990).
Summary
Cappetta & Nolf 2005 is a useful and valuable resource for organizing and identifying Paleogene sand tiger teeth. From a European perspective, it provides a good history and clarifies many matters such as Orpodon, Carcharias sp., places robustus within Jaekelotodus and removes Carcharias atlasi from the European faunal list.
The primary fault to be found is that genera (as argued) were erected on the basis of 'tooth-design' only. Many of these taxa are known from thousands of teeth and the authors appear to have made no effort to reconstruct the tooth-sets and use 'dentition-design' as the basis for erecting taxa. Although reasonable, their conclusions would have been better served by presenting examples of tooth-positions A1-L3 (both upper & lower) rather than the abbreviated text and illustrations included. The authors do indicate that this is only the initial step which will be followed by a revision of the Odontaspididae from Morocco and other African countries; they may intend to provide greater evidence in later papers.
This review may appear harsh to some, but we are dealing with a very important topic presented by a (arguably the) preeminent authority on fossil elasmobranch teeth.
Acknowledgements
We would like to thank the others who have assisted in this project by providing specimens, literature, and/or insights, as well as those that took the time to review and comment upon the content; they include: Richard Chandler, Gilles Cuny, Jacques Herman, Gino Marien, Fabrice Moreau, Gary Grimsley, Gordon Hubbell, Guy Van den Eeckhaut, David Ward and Pat Young.
Footnotes
| 1. |
Genera not represented in the C&N North Sea basin were listed but not fully discussed in the C&N paper and are subsequently not being included in this webpage. |
| 2. |
Not included in Cappetta & Nolf but added as supplementary reference for this webpage. |
| 3. |
Authors of this webpage are of differing opinions as to whether or not stunted anteriors or intermediate spacer teeth should be included in the tooth-set reconstructions. One author (SC) argues:
These small teeth are subject to collecting-bias and are therefore not sufficiently represented to tender statistical evidence.
By their very nature, they lack sufficient diagnostic similarity with their significant counterparts to comfortably establish a relationship.
The asterisk indicates that teeth are present in the samples that could be ascribed to these positions but not with 100% certainty. |
Selected References
Agassiz, L. J. R. 1833-1844. Recherches sur les poissons fossiles. Text (5 vols; I., xlix+188 pp., II xii+310+366 pp., III viii+390 pp., IV xvi+296 pp., V xii+122+160 pp.) and Atlas (5 vols; I 10 pl., II., 149 pl., III 83 pl., IV, 61 pl., V, 91 pl.). Neuchâtel.
Cappetta, H & Nolf, D, 2005. Revision de quelques Odontaspidae (Neoselachii: Lamniformes) du Paleocene et de l'Eocene du Bassin de la mer du Nord Bulletin de l'institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre 75:237-266.
Cappetta, H. 1987. Chondrichthyes II. Mesozoic and Cenozoic Elasmobranchii. In: Handbook of Paleoichthyologie, vol. 3b, Gustav Fischer Verleg, Stuttgart, 193 pp.
Casier, E., 1967. Le Landénien de Dormaal (Brabant) et sa faune ichthyologique. Mémoires de l'Institut royal des Sciences naturelles de Belgique, 156: 1-66.
Cunningham, S. B. 2000. A comparison of isolated teeth of early Eocene Striatolamia macrota (Chondrichthyes, Lamniformes), with those of a Recent sand shark, Carcharias taurus. Tertiary Research 20(1-4): 17-31.
Glikman, L., 1964. Akuly paleogena i ikh stratigraficheskoe znachenie Akademii Nauk Soyuza Sovetskikh Sotsialisticheskikh Respublik, 1-228 Moscow.
Herman, J., 1972. Les vertébrés du Landénien inférieur (LIa ou Heersien) de Maret (Hameau d'Orp-Le-Grand). Bulletin de la Société belge de Géologie, de Paléontologie et d'Hydrologie, 81 (3-4): 191-207.
Jaekel, O., 1895. Unter-Tertiäre Selachier aus Südrussland. Mémoires du Comité Géologique de Saint-Pétersbourg, 9: 19-35.
Leriche, M. 1905. Les Poissons Paléocènes de la Belgique. Mémoires du Musée Royale d'Histoire Naturelle de Belgique. Bruxelles. 33: 49-228.
Leriche, M., 1921. Sur les restes de poissons remaniés dans le Néogène de la Belgique. Leur signification au point de vue de l'histoire géologique de la Belgique pendant le tertiaire supérieur. Bulletin de la Société belge de Géologie, 30: 115-120.
Kent, B. 1994. Fossil Sharks of the Chesapeake Region. 146 pp. Egan Rees & Boyer, Maryland.
Kent, B. 1999. Sharks from the Fisher/Sullivan Site. In: Early Eocene Vertebrates and Plants from the Fisher/Sullivan Site (Nanjemoy Formation) Stafford County, Virginia, Weems, R. & Grimsley, G., 1999.
Virginia Division of Mineral Resources, Pub 152. pp 11-38.
Menner, V. V. 1928. The Palaeogene sharks of Mangyschlak, Emba and from the east of Oural. Bulletin de la Société des Naturalistes de Moscou Section, Géologique vol. 6(3-4): 291-338. Moscow In Russian.
Moreau, F. & Mathis, S. 2000. Les élasmobranches du Thanétien (Paléogène) du nord de la France, des carrières de Templeuve et de Leforest. Cossmanniana.
Nolf, D., 1988. Fossiles de Belgique. Dents de requins et de raies du Tertiaire de la Belgique. Institut Royal des Sciences Naturelles de Belgique, 184 pp.
Rafinesque, C. S. and [As Rafinesque Schmaltz, C. S.] 1810. Caratteri di alcuni nuovi generi e nuove specie di animali e piante della sicilia, con varie osservazioni sopra i medisimi. (Part 1 involves fishes, pp. [i-iv] 3-69 [70 blank], Part 2 with slightly different title, pp. ia-iva + 71-105 [106 blank]). Caratteri.
Siverson, M., 1992. Biology, dental morphology and taxonomy of Lamniform sharks from the Campanian of the Kristianstad basin, Sweden. Palaeontology, 35 (3): 519-554.
Siverson, M., 1995. Revision of the Danian Cow Sharks, Sand Tiger Sharks, and Goblin Sharks (Hexanchidae, Odontaspididae, and Mitsukurinidae) from Southern Sweden, Journal of Vertebrate Paleontology, 15 (1): 1-12.
Ward, D. J. 1988. Hypotodus verticalis (Agassiz, 1843), Hypotodus robustus (Leriche, 1921) and Hypotodus heinzelini (Casier, 1967), Chondrichthyes, Lamnifirmes, Junior Synonyms of Carcharias hopei (Agassiz, 1843). Tertiary Research, 10: 1-12.
White, E.I., 1931. The vertebrate faunas of the English Eocene. I. From the Thanet Sands to the Basement Bed of the London Clay. British Museum (Natural History), 121 pp.
Winkler, T.C., 1874a. Mémoire sur quelques restes de Poissons du Système Heersien.In:
Archives du Musée Teyler, 1878; vol. IV, (fasc. 1, 1876), pp 1-15; Extraits [1874]. Les Héritiers Loosjes, Haarlem, Belgium.
Winkler, T.C., 1874b. Deuxième mémoire sur des dents de poissons fossiles du terrain bruxellien. In: Archives du Musée Teyler, 1878; vol. IV (fasc. 1, 1876), pp 16-48; Extrait, 10 pages. Les Héritiers Loosjes, Haarlem, Belgium.
Zhelezko , V. I. & Kozlov, V. A. 1999. Elasmobranchii and Palaeogene biostratigraphy of Transurals and Central Asia. Materials on stratigraphy and Palaeontology of the Urals Vol. 3. Russian Academy of Sciences Urals Branch Uralian Regional Interdepartment Stratigraphical Comissian, Ekkaterinburg. 324 pp, 61pls.
| |
{kind=link}