| |
Participation was certainly optional, but when the concerned citizens of Nanjemoy were meeting to discuss the BLM's involvement in the management of Potomac River lands that contain Aquia exposures, even a New Yorker's interests were piqued — it seemed relevant. So three of us (JB, GG & BH) found ourselves listening to the land management discussion. During a break, Gary turned to me (JB) and remarked that a website highlighting the Aquia fauna could be a useful tool for those arguing that public access should be permitted. Despite my reservations to starting a new project, I turned to Bill to get his opinion. It was March 2nd and we agreed to invest the time that would be required to fully research the topic and present a webpage. Rob would later join the project when stratigraphic insight was required. In looking over the final product, Gary's suggestion led far beyond our initial objectives. The presented material builds upon the foundation laid down by Ward & Wiest (1990) and will hopefully serve as a starting point for others.
Best known from cliffs exposed along the Potomac River (Virginia & Maryland) and for the teeth of Palaeocarcharodon, the Aquia has been popular to generations of collectors and paleontologists. Although the surface-collecting of beach material is most common, the faunal list as presented is based on in situ material (unless noted to the contrary). Sampling for this presentation has been largely limited to the Piscataway and lower sediments of the Paspotansa Members. Over time, upper Paspotansa observations will be incorporated on this page.
Stratigraphy
From New Jersey to Georgia, much of the Atlantic Coastal Plain lies above Late Palaeocene sediments. The various formations correspond with the embayments of the paleo-coast: Vincentown (Raritan), Aquia (Salisbury), Beaufort (Albemarle) and Black Mingo Formation (Charleston Embayment). The Southeast Georgia Embayment is represented by the Tuscahoma Frm., Nanafalia Frm. and Porters Creek Clay. In Virginia and Maryland, the Aquia is not only exposed along the Potomac, but by various creeks, rivers (Pamunkey & Rappahannock) and construction sites.
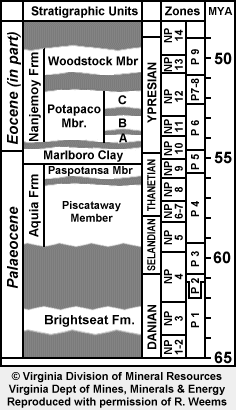 The Paleogene stratigraphy of the Salisbury Embayment has been studied and redefined continually since the nineteenth century. Ward (1985) attempted to synthesize these studies, and included in the Pamunkey Group: Brightseat Frm. (Lwr Palaeocene), Aquia Frm. & Marlboro Clay (Upper Palaeocene), Nanjemoy Frm. (Lwr Eocene), Piney Point Frm. (Mid Eocene) and Chickahominy Frm. (Upr Eocene). According to Gibson, et al (1991), in most locations, the Aquia unconformably overlies Cretaceous units; however, in a limited area east and southeast of DC, it unconformably overlies the Brightseat Formation.
The Paleogene stratigraphy of the Salisbury Embayment has been studied and redefined continually since the nineteenth century. Ward (1985) attempted to synthesize these studies, and included in the Pamunkey Group: Brightseat Frm. (Lwr Palaeocene), Aquia Frm. & Marlboro Clay (Upper Palaeocene), Nanjemoy Frm. (Lwr Eocene), Piney Point Frm. (Mid Eocene) and Chickahominy Frm. (Upr Eocene). According to Gibson, et al (1991), in most locations, the Aquia unconformably overlies Cretaceous units; however, in a limited area east and southeast of DC, it unconformably overlies the Brightseat Formation.
The Aquia is made up of clayey, silty and shelly glauconitic sands and has been divided into 'zones' (levels/beds) which were ascribed to either the Piscataway (lower) or Paspotansa (upper) Members. Clark and Martin (1901) had attributed 'zones' 8 and 9 to the Paspotansa and the balance to the Piscataway. Ward (1985) noted that the poorly sorted sands of 'zones' 1-5 stood in contrast to the well sorted sands of 'zones' 6 & 7, and that the molluscan fauna coincidentally changed across this boundary — he revised the Paspotansa to include 'zones' 6 and 7. To the casual collector, it is the abundance of Turrittella shells that alerts one to these Paspotansa sediments.
The clayey sands of 'zone' 1 (Piscataway) have limited exposures in Maryland and Virginia. More commonly exposed and collected is 'zone' 2. These shelly sands are quite rich in vertebrate material and appear to be the primary source of Palaeocarcharodon teeth. 'Zone' 3 is an indurated ledge (limestone appearance, and when present/observable) clearly demarking the top of 'zone' 2 and bottom of 'zone' 4. 'Zone' 4 appears to be quite variable depending on depositional location (?depth). Along the Potomac (Charles County), it is a shelly sand (similar to 'zone' 2) and somewhat thick (see below image). Further inland (Prince Georges County), shell content and layer thickness diminishes. A second ledge ('zone' 5) is the upper limit of the Piscataway based on Ward (1985).
 |
| Piscataway 'zones' 2-5, Charles County, MD |
The accompanying stratigraphic chart follows Berggren et al. (1995) which splits the Palaeocene as follows: Danian (65.0 Ma to 61.0 Ma), Selandian (61.0 Ma to 57.9 Ma) and Thanetian (57.9 Ma to 54.8 Ma). Thus the Selandian is now what used to be the lower part of the Thanetian. The Aquia starts in upper NP 5, which is late Selandian, and runs through most (but not all) of the Thanetian. Because most authors have continued to employ Thanetian for the period 61.0 to 54.8 Ma, this webpage and the associated genus pages do not reflect this renaming (be it justified or not).
Depositional Environment
In Charles County, the Piscataway depositional environment is shallow marine, perhaps one to ten miles offshore. The very abundant plant debris and the occasional mammal bones and teeth indicate that land was not too terribly far away. Probably at that time, the shoreline lay roughly along the corridor of Interstate 95. Understandably, Prince Georges County marine fossils suggest shallower waters.
The three kinds of crocodiles indicate that the climate was much warmer than present, perhaps like south Florida, though not quite as warm as in the early Eocene. The pine cones suggest that there probably were barrier islands covered with pine forests. Most kinds of pollen were from extinct trees, but birch (Betula), alder (Alnus), and hickory (Carya) pollen have been reported. Piscataway sediments are fine to medium grained, massively bedded due to intense bioturbation, and usually abundantly glauconitic. The abundance of mollusk shells and occasional limestone-cemented beds indicate slow deposition.
Partial skeletons of crocs and turtles are fairly abundant, but almost none of them are very complete. The three mammal occurrences described by Ken Rose (2000), along with Planetochelys (a primitive box turtle) indicate at least some proximity to shore or an offshore river current bringing in carcasses. The Paspotansa appears to have been a much more offshore deposit, as wood is not very abundant and the sediments better sorted.
Chondrichthyan Fauna
Scarcity:
Rare - One or two specimens found each year (by all collectors).
Scarce - A regular collector may find one or two a year.
Uncommon - A collector may find one every few trips.
Common - A specimen can be found each trip.
Abundant - Pretty hard not to find a number of specimens each trip.
|
|
|
|
|
CLICK ON IMAGES FOR GENUS INFORMATION
|
 — Indicates that species not linked to a genus page.
— Indicates that species not linked to a genus page.
|
 |
|
Ischyodus dolloi - Ratfish
Ratfish material (two or three species) such as this left mandibular
is present but uncommon in the Aquia.
| |
|
SYNECHODONTIFORMES Duffin & Ward 1993 |
 |
|
Paraorthacodus clarkii
EASTMAN, 1901 - Extinct shark.
Uncommon in the Piscataway. Specimens have been recovered directly from
'zones' 1, 2 and 6/7. | |
|
HEXANCHIFORMES Buen 1926 |
|
|
|
It is uncertain at this time which genera
might be present in the fauna. Kent (1994) reported Notorynchus serratissimus
(AGASSIZ, 1843) however the authors have been unable to
confirm this observation. Two specimens, thought to represent Notidanodon
(paper in process), have also been reported. Needless to say, these are rare (if
present) in the fauna. |
|
 |
|
cf Hexanchus sp - Extinct Sixgilled shark.
Not previously reported from the Aquia, this upper tooth was found
in situ ('zone' 7) and compares well with those of Hexanchus.
|
|
SQUALIFORMES Goodrich 1909 |
 |
|
Squalus cf minor
(WINKLER, 1874) - Dogfish shark.
Teeth of this design appear to be relatively common, at least in
'zone' 2 of the Aquia. |
 |
|
"Megasqualus" orpiensis
WINKLER, 1874 - Extinct dogfish shark genus.
Appears to be somewhat uncommon in 'zone' 2 Cappetta has questioned
the validity of the genus Megasqualus.
|
|
SQUATINIFORMES Buen 1926 |
 |
|
Squatina "prima"
(WINKLER, 1874) - Angel shark.
Teeth of this design are generally uncommon in Aquia sediments, although they
appear more common in 'zone' 2.
|
|
HETERODONTIFORMES Berg 1937 |
 |
|
Heterodontus cf lerichei
CASIER 1943 - Bullhead shark.
Although uncommon in Aquia sediments along the Potomac, teeth from this
species are common in 'zone' 2 deposits in Prince Georges Co. . |
|
ORECTOLOBIFORMES Applegate 1972 |
 |
|
Ginglymostoma cf subafricanum
ARAMBOURG 1952 - Nurse shark.
Uncommon to scarce in Piscataway sediments.
|
 |
|
Delpitoscyllium africanum
(LERICHE 1927) - Extinct nurse shark.
Uncommon to scarce in Piscataway sediments. |
|
|
|
Palaeorhincodon wardi
HERMAN, 1975 - Extinct whale shark.
Kent (1994) listed this species as present in the fauna, but these authors
have been unable to confirm this observation. David Ward notes (pers com 6-15-02)
that wardi is an Eocene species, and if present these teeth would likely
be dartevelli.
|
|
LAMNIFORMES Berg 1958 |
| In 2005, Cappetta and Nolf released Revision de quelques Odontaspidae (Neoselachii: Lamniformes) du Paleocene et de l'Eocene du Bassin de la mer du Nord. The impact of this paper greatly effects the naming of many of the North American taxa as well. In order to judge it's relevancy and allow others to evaluate their determinations, elasmo.com provides a review of this paper. |
| |
 |
|
Anomotodon cf novus
(WINKLER 1874) - Extinct goblin shark genus.
Teeth from this species are common in the Aquia. |
 |
|
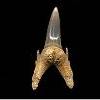
Odontaspis winkleri
LERICHE, 1905 - Smalltooth sand tiger.
A common species in Piscataway sediments. |
 |
|
Palaeohypotodus rutoti
(AGASSIZ, 1843) - Extinct sand tiger genus.
Uncommon in Aquia sediments.
|
 |
|
Brachycarcharias lerichei
(CASIER, 1946) - Extinct sand tiger genus.
Common in Aquia sediments, previously attributed to Serratolamna. |
 |
|
Hypotodus verticalis
(AGASSIZ, 1843) - Extinct sand tiger genus
A common tooth in Aquia sediments, previously included as Carcharias hopei. |
 |
|
Striatolamia striata
(WINKLER, 1874) - Extinct sandtiger- like shark genus.
The shark that ate the Paleogene — teeth are abundant in the Aquia. |
 |
|
Striatolamia sp
- Extinct sandtiger- like shark genus.
This tooth-design is common in Piscataway sediments.
|
 |
|
Jaekelotodus robustus
(LERICHE, 1921) - Extinct lamniform genus
Formerly listed as Carcharias robustus, the accompanying specimen is
from 'zone' 2 float.
|
 |
|
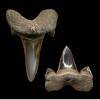
Isurolamna inflata
(LERICHE, 1905) - Extinct mackerel shark genus.
Somewhat uncommon in Aquia sediments. Illustrated teeth from 'zone' 2. Crown is smooth,
more I. affinis-like in design. |
 |
|
Cretalamna appendiculata
(AGASSIZ 1843) - Extinct mackerel shark genus.
Common in Aquia sediments.
|
 |
|
Otodus obliquus
AGASSIZ, 1843 - Extinct mackerel shark genus.
Common in Aquia sediments, accompanying specimen from Piscataway of Virginia.
|
 |
|
Palaeocarcharodon orientalis
(SINZOW, 1899) - Extinct lamniform genus.
Relatively uncommon in Aquia sediments, specimens are derived from 'zone' 2, either
in situ or in float. These teeth have been reported from float thought to be Paspotansa, but the actually origin has yet to be proven. |
|
CARCHARINIFORMES Compagno 1973 |
 |
|
Pachygaleus lefevrei
(DAIMERIES, 1891) - Extinct houndshark genus.
Teeth of this species are relatively common in Piscataway sediments. |
 |
|
Galeorhinus sp. - Extinct Tope Shark
The Aquia yields a number of Galeorhinus-like tooth-designs. The accompanying
specimen shows similarities
with G. mesetaensis NOUBHANI & CAPPETTA (1997)
a Palaeocene species from Morocco. |
 |
|
Palaeogaleus sp
- Extinct Houndshark
Uncommon in studied Piscataway sediments. Noubhani (pers. comm. 07-25-02):
By its size and general morphology, it is similar to P. cf larachei
NOUBHANI & CAPPETTA (1997), from the Thanetian of
Morocco.
|
 |
|
Triakis sp
- Extinct Houndshark
Compares with T. antunesi NOUBHANI &
CAPPETTA 1997 (Palaeocene, Morocco).
Ward (pers. comm. 5-20-02) indicated that this design was listed
as S. brivesi in Ward & Wiest (1990).
Noubhani (07-25-2002): Undoubtedly Triakis species, but doesn't
compare well with the Moroccan ones.
|
 |
|
? Scyliorhinus sp - Catshark.
Uncommon. Labial face of crown is smooth, accompanying specimen
from Paspotansa. According to Ward (pers. comm. 5-20-02), reported as
Scyliorhinus gilberti CASIER, 1946 by
Ward & Weist (1990). Noubhani (pers. comm. 7-25-2002) "could be a
Scyliorhinus species." |
 |
|
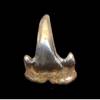
Scyliorhinus cf ptychtus
NOUBHANI & CAPPETTA 1997 - Extinct Catshark.
Reported by N&C (1997) as from Palaeocene & Ypresian
sediments of Morocco. Accompanying image from beach below Piscataway exposures. |
 |
|
Premontreia cf subulidens
(ARAMBOURG, 1952) - Extinct catshark genus.
Uncommon the Piscataway. Noubhani & Cappetta (1997) reported species
from the Paleogene of Morocco. Kent (1999)
attributed Eocene teeth of this design to P. degremonti
CAPPETTA, 1992. Imaged tooth from Piscataway beach, another
damaged specimen in situ 'zone' 2. |
 |
|
Abdounia beaugei (ARAMBOURG, 1935) - Extinct requiem shark genus.
Relatively scarce in the Piscataway, these teeth are more common in Paspotansa sediments. |
|
|
|
Abdounia sp - Extinct requiem shark genus.
Reported by Ward & Weist (1990). |
|
RAJIFORMES Berg 1940 |
|
|
|
A single specimen was reported by Ward & Weist (1990) as possibly a skate.
Ward (pers. com. 2002) indicated that this may have been a smooth-crowned
dasyatid. For now, the presence of "Raja" in the fauna is questionable.
|
|
MYLIOBATIFORMES Compagno 1973 |
 |
|
Hypolophodon cf sylvestris - Extinct stingray genus.
Common in Piscataway sediments. |
 |
|
Coupatezia woutersi
(WINKLER, 1874) - Extinct stingray genus
This species was reported by Ward & Weist (1990) from the Piscataway. The accompanying
(damaged) specimen is the only 'zone' 2 example collected by this page's authors.
More common in the Paspotansa. |
 |
|
Dasyatis sp
- Whiptail Ray
The accompanying specimen is from 'zone' 2. Dasyatis teeth are uncommon in the Piscatawy. |
 |
|
cf "Myliobatis" dixoni AGASSIZ, 1843 - Eagle ray genus
Common in Piscataway sediments. Isolated teeth found in situ, accompanying image from 'zone' 2 float. |
 |
|
"Myliobatis" sp - Eagle ray genus
Common in Piscataway sediments. Isolated teeth found in situ, accompanying image from 'zone' 2 float. |
|
|
|
Other myliobatids
Isolated myliobatid teeth are abundant in the Piscataway.
However, unless these teeth have diagnostic characteristics
or are part of complete tooth plates, identifications are questionable.
More species may be represented than listed above.
|
 |
|
Undescribed myliobatoid 1
- Eagle ray genus
Known from isolated teeth only ('zone' 2 and beach). These teeth were originally thought to be "M." nzadinrnsis-like in design. However, based on diagnostic characteristics, it is difficult to determine if these teeth should be placed with the myliobatids or rhinopterids.
|
 |
|
Rhinoptera sp - Cow-nose ray genus
Common in Piscataway sediments, teeth may represent more than one species. |
 |
|
"Myliobatis" sulcidens - Extinct ray species
Relatively uncommon and usually as fragments only in the lower Aquia. |
 |
|
Burnhamia sp - Extinct mobulid ray genus
Ward & Weist (1990) reported this genus from the Paspotansa only. The
accompanying specimen is from the Piscataway 'zone 2' and should be considered
uncommon to scarce in the Piscataway. |
Footnotes
| 1. |
| Ward (1995) ascribed the common molluscan fauna as follows: |
| Piscataway |
Paspotansa |
Cucullae gigantea CONRAD 1830
Ostrea alepidota DALL 1898
Crassatellites capricranium (ROGERS 1839)
Lucina aquiana CLARK 1895
Dosiniopsis lenticularis (ROGERS 1839)
Pitar pyga CONRAD 1845
Corbula aldrichi MEYER, 1885
Turritella humerosa CONRAD 1835
Turritella mortoni CONRAD 1830
|
Cucullae gigantea CONRAD 1830
Pycnodonte sp
Ostrea sinuosa ROGERS & ROGERS 1837
Crassatellites alaeformis CONRAD 1830
Venericardia regia CONRAD 1865
Pitar pyga CONRAD 1845
Turritella humerosa CONRAD 1835
Turritella mortoni CONRAD 1830
Lunatia marylandica CONRAD 1865
| |
| 2. |
The origin of these hard beds has been of long debate. They do seem to be diagenetic, but the question is whether they are early diagenesis or late diagenesis. If early, they are pretty much parallel to original stratification. If late, they may follow groundwater tables and thus may well cross original stratification boundaries. There appears to be no convincing evidence that they are hardgrounds, which would represent old sea bottoms. There don't appear to be any phosphate crusts, borings, or oyster banks that would be expected from hardgrounds. The hardbeds could, however, be old coquina deposits that had their interstitial spaces filled early-on with carbonate cement. It could represent a rapidly filled shallow shelf environment that had fairly strong currents sweeping through. |
| 3. |
Eosuchus lerichei Dollo, 1907, Thoracosaurus neocesariensis (de Kay, 1842) and an undescribed species. All three are long-snouted forms, so the third is not an alligatorine form like Bottosaurus or Allognathosuchus. |
Acknowledgements
Although four names may appear on the top of this webpage, its creation extends well beyond our
time and efforts. Others who participated in this cyber-venture (intentionally or not) include
Ron Blakey, Steve Cunningham, Abdelmajid Noubhani, Daryl Serafin, David Ward and David Wells. We sincerely appreciate their assistance.
Berggren, W.A., Kent, D.V., Swisher, C.C. III, and Aubry, M.-P., 1995, A revised Cenozoic geochronology and chronostratigraphy: in Geochronology, Time Scales, and Global Stratigraphic Correlation, SEPM Special Publication #54, p. 129-212.
Cappetta, H & Nolf, D, 2005. Revision de quelques Odontaspidae (Neoselachii: Lamniformes) du Paleocene et de l'Eocene du Bassin de la mer du Nord Bulletin de l'institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre 75:237-266.
Clark, W.B. & Martin, G.C., 1901, The Eocene deposits of Maryland. Maryland Geological Survey, Eocene volume, p1-92, 122-204.
Gibson, T.G., Bybell, L.M., and Govoni, D.L., 1991, Paleocene and Eocene strata of the central Atlantic Coastal Plain, p. 1-14, in Gibson, T.G., and Bybell, L.M., eds., Paleocene-Eocene boundary sedimentation in the Potomac River Valley, Virginia and Maryland: International Geological Correlation Programme Project 308, Field Trip Guidebook, 124 pp.
Kent, B. W. 1994. Fossil Sharks of the Chesapeake Region. Egan Rees & Boyer, Maryland. 146 pp.
Rose, K.D., 2000. Land-mammals from the Late Paleocene Aquia Formation: The first early Cenozoic mammals of Maryland, in: Proceedings of the Bio. Scty. of Washington, 113(4):855-863.
Ward, D.J. & Wiest, R.L., 1990. A checklist of Palaeocene and Eocene sharks and rays (Chondrichthyes) from the Pamunkey Group, Maryland and Virginia, USA. Tertiary Res., 12(2) p 81-88.
Ward, L.W., 1985. Stratigraphy and Characteristic Mollusks of the Pamunkey Group (Lower Tertiary) and the Old Church Formation of the Chesapeake Group — Virginia Coastal Plain. U.S. Geo. Srvy. Prof Paper 1346, 78 pp.
Weems, R.E. & Grimsley, G.J., 1999. Introduction, Geology, and Paleogeographic Setting in: Early Eocene Vertebrates and Plants from the Fisher/Sullivan Site (Nanjemoy Formation) Stafford County, Virginia. Virginia Division of Mineral Resources, Pub 152. pp 1-10.
|